A nem-strukturális fehérjék funkcióinak áttekintése
2014.03.28. 16:01
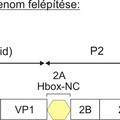
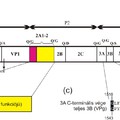
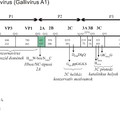
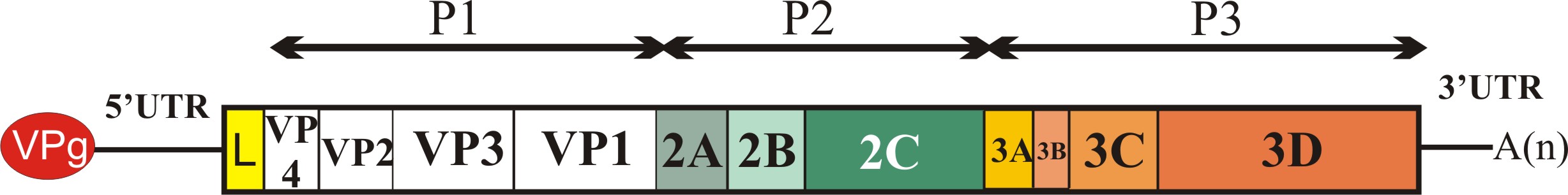
Az úgynevezett nem-strukturális fehérjék csoportjába a picornavírus genom P2 és P3 régióján (lásd ”A picornavírus genom felépítése” bejegyzést) valamint egyes vírusoknál a P1 régió előtti szakaszon (Leader fehérje kódoló szakasz) kódolt virális fehérjék tartoznak. A csoportot különböző (olykor összetett) funkcióval rendelkező fehérjék alkotják. Ebben a részben megpróbálom röviden áttekinteni az egyes ismert nem-strukturális fehérjék funkcióit, az esetleges funkciók nagy száma miatt a teljesség igénye nélkül. Az egyes fehérjéket a vírus genomon található elhelyezkedésük (lásd a bejegyzés ábráját) sorrendjében tárgyalom.
Leader
Minden olyan fehérjét Leader-nek neveznek, amely a picornavírus genom P1 régiója (kapszid-fehérjék) előtti részen kódolódik függetlenül attól, hogy az milyen funkcióval rendelkezik. Az eddig ismert emlős picornavírusok több mint fele kódol valamilyen Leader fehérjét. Az egyes Leader fehérjék mérete nagyon változékony, az egészen rövidtől (pár tíz aminosav) akár 462 aminosav hosszúságig változhat. Az eddig azonosított Leader fehérjék jelentős részéről nem tudjuk, hogy milyen funkciót lát el a vírus replikációjában. Az FMDV (papain-szerű cisztein proteáz) és a cardiovírusok (Zn-kötő fehérje) Leader fehérjéi szerepet játszanak a virális poliprotein érési folyamataiban, a gazdasejt CAP-függő transzlációjának leállításában, valamint a gazdaszervezet immunfolyamatainak kijátszásában.
2A
A virális kapszid fehérjéket (VP4, VP2, VP3 és VP1) kódoló régió (P1) után funkcionális szempontból igen változatos fehérjéket kódoló régiót találunk. A 2A régió az ismert emlős picornavírusok egy kis részében (pl. Ljungan vírus, Pasivirus, vagy a tengeri emlősöket fertőző aquamavírusok) nem egy, hanem két fehérjét (2A1 és 2A2) kódol (ún. dicisztronos 2A régió), azonban az emlős picornavírusok döntő többségében csak egy fehérjét kódoló, ún. monocisztronos 2A genom régiót találunk.
Jelen ismereteink szerint összesen 4 különböző 2A típust ismerünk:
tripszin-proteáz funkciójú 2A (pl. az Enterovírus és valószínűleg a Sapelovírus nemzetségekbe tartozó picornavírusokban), amely képes saját magát leválasztani a virális poliproteinről, emellett több, a gazdasejt sejtplazmájában előforduló, a gazdasejt ún. CAP-függő fehérjeszintéziséhez nélkülözhetetlen transzlációs faktorokat (pl. eIF-4G) is képes elbontani.
Aphthovírus-szerű 2A fehérjék változatos hosszúságúak (20 - 145 aminosav) és valószínűleg változatos funkcióval is rendelkeznek. Ilyen típusú 2A fehérjét találunk egyes emlős picornavírusok, mint például az Aphthovirus, Cardiovirus, Erbovirus, Teschovirus, és Senecavirus nemzetség tagjai között.Ebbe a csoportba tartozó 2A fehérjék mindegyike ugyanarra a konzervatív aminosav-motívumra (DxExNPGP, ahol x bármilyen aminosav lehet) végződik. Ezt az aminosav-motívumot ”riboszóma-ugró” motívumnak is nevezik. A virális genom transzlációja során, a genomon végighaladó riboszómák (80S) elérik a DxExNPG↓P (↓: a ”hasadási helyet” jelöli) motívumot akkor a motívum utolsó két aminosava (G és P) között nem alakítanak ki peptidkötést, ami következtében az NPG-re végződő virális polipeptid darab leválik a riboszómáról, miközben a riboszóma folytatja a genom hátralevő részéről a fehérje szintézist. Emellett egyes aphthovírus-szerű 2A fehérjék (pl. cardiovírusok) szintén részt vesznek a gazdasejt CAP-függő fehérjeszintézisének leállításában.
Hepatitisz A-szerű 2A fehérjét egyelőre csak a Hepatitis A vírusokban írtak le. A 2A fehérjének nincs ismert, az előző két fehérjéhez hasonló fehérje-hasító funkciója. A 2A egy része a VP1 kapszid fehérjékhez kötötten (pX) jelenik meg a virion felszínén, szerepe lehet a virionok gazdasejt-membránnal történő beburkolódásának folyamatában (További olvasmány a témában itt)
Hbox-NC 2A fehérjék szintén elterjedtek az emlős picornavírusok körében. Ilyen típusú 2A fehérjét kódolnak az Aquamavirus, Kobuvirus, Parechovirus, és ”Rosavirus” nemzetség tagjai. A csoport 2A fehérjéi jelentős szerkezeti és szekvencia-rokonságot mutatnak az eukarióta H-rev107 fehérjékkel, melyek fontos sejt-proliferáció szabályozó szereppel rendelkeznek.
2B
A picornavírusok 2B proteinjeinek mérete viszonylag tág határok között mozog: 98 aminosav (rhinovírusok) és 283 aminosav (Equine rhinitis B virus). A 2C virális peptiddel alkotott fúziós fehérje (2BC) részeként részt vesz sejtmembrán-vezikulák kialakításában, amelyen picornavírus RNS genom replikációja zajlik. Az enterovírusok szabad 2B fehérjéi membrán-pórusokat formálhatnak a sejt Golgi apparátus, illetve endoplazmatikus retikulum membránjain, azonban ez az úgynevezett viroporin-funkció más picornavírusoknál (pl. aphtho-, cardio-, vagy hepatovirusoknál) nem azonosítható. A 2B fehérje funkciója a legtöbb picornavírusnál jelenleg ismeretlen.
2Chel
A 2C egy multifunkcionális vírusfehérje, amelynek szerepe van többek közt a vírus replikációjában illetve sejtmembrán-vezikulák kialakításában. Minden eddig ismert 2C fehérje rendelkezik egy ATP-kötő/bontó doménnel, ami az ún. III. helikáz szupercsaládba (HELSF3) tartozik. Hasonló HELSF3 doménnel rendelkező virális enzimek igen elterjedtek a vírusok világában. A 2C fehérjék vége általában egy amfipatikus szakaszt tartalmaz, amelynek szerepe lehet a 2C membránhoz kötődésében.
3A
Hasonlóan a 2B fehérjékhez a 3A peptidek mérete is igen változó: 76 (rhinovírus) és 150 (FMDV) aminosav között változhat. FMDV esetében a 3A hossza, a vírus fertőzőképessége és gazdafaj-spektruma között szoros kapcsolat áll fent. 3A-nak illetve 3B-vel alkotott fúziós fehérjének (3AB) fontos szerepe van a virális genomról történő fehérjeszintézis megindításában, illetve a 3D virális fehérje (lásd alább) ko-faktoraként is funkcionálhat, azonban ezek a feltételezett funkciók nem teljesen elfogadottak. Egyes enterovirális 3B proteinek képesek az endoplazmatikus retikulum és a Golgi apparátus közti fehérje-transzportot blokkolni. Azonban ez a 3B funkció sem univerzális a picornavírusok között.
3B
A 3B genom régió által kódolt rövid (21-23 aminosav) fehérje (VPg, 3BVPg) kovalensen kötődik minden ismert picornavírus RNS genomjának 5’ végéhez. A kötés kulcs aminosavja a VPg 3. tirozinja, ami minden ismert picornavírus VPg-ben azonos. A kisméretű VPg hidrofób részeket tartalmaz és erősen bázikus tulajdonságú aminosavakból áll. Egyes vírusnemzetségek tagjainál a VPg-kódoló 3B régió duplikált (mosavírusok, aquamavírusok) vagy akár triplikált (FMDV) formában is jelen lehet a vírusgenomon. A VPg fehérje védi a virális RNS-t a gazdasejt RNÁzaitól és fontos szerepe van a virális RNS genom negatív szálának szintézisében is.
3Cpro
A viszonylag nagyméretű (180-220 aminosav), cisztein-proteáz funkciójú 3C fehérje végzi el a virális poliprotein érési folyamata alatt lejátszódó hasításának dandárját. Emellett érett formában (3C) vagy 3D-vel alkotott fúziós fehérjeként (3CD) több gazdasejt sejtplazmájában előforduló fehérjék, transzkripciós és transzlációs faktorok inaktiválásáért is felelős. Ezáltal leállítja a gazdasejt transzkripciós és CAP-függő transzlációs folyamatait. Emellett minden ismert 3C fehérje rendelkezik egy erősen konzervatív RNS-kötő régióval is. A 3C-szerű cisztein proeteázok elterjedtek a pozitív egyszálú RNS vírusok körében is.
3DRdRp
A picornavírusok legkonzervatívabb fehérjéje a 3D, amely egy nagyméretű (460-470 aminosav) RNS-függő RNS polimeráz (RNA-dependent RNA polymerase, RdRp) funkciójú enzim. Ez a virális enzim képes mind a negatív, mind a pozitív RNS szál szintézisére a virális replikáció alatt. A 3D fehérje rokonságot mutat számos, replikáz funkciójú enzimmel, pl. DNS-, illetve RNS-függő DNS polimerázokkal.