

A picornavírusok 5’ nem kódoló régióinak jellemzése
2014.05.12. 10:14
A picornavírusok (és egyéb más pozitív egyszálú RNS vírusok, pl. Hepatitisz C - Flaviviridae család) 5’ nem kódoló régiója (5’ UTR = UnTranslated Region) komplex másodlagos struktúrával rendelkezik, amely funkcionális szempontból két különböző régióra osztható. Az 5’ vég olyan strukturális elemeket tartalmaz, amelyek nélkülözhetetlenek a virális genom megsokszorozódásához. A transzláció iniciációs kodon (start kodon) előtti szakasz az ún. belső riboszóma kötőhely (IRES=Internal Ribosomal Entry Site) a virális poliprotein képződésének megindításáért felelős régió, amely a riboszómális komplex vírusgenomhoz történő kapcsolódását biztosítja. Régóta ismert, hogy az eukarióta mRNS molekulák transzlációja úgynevezett CAP-függő folyamat. (CAP azaz ”sapka” az érett eukarióta mRNS molekulák 5’ végéhez kovalensen kötött 7-metil-guanozin trifoszfát molekula, amely egyrészt védi az mRNS-t az RNS bontó enzimektől, másrészt transzláció iniciációs faktor-kötőhelyként szolgál. Bővebb információ az alábbi linken). Az eukarióta mRNS molekulákkal ellentétben a picornavírusok (és egyes egyéb más pozitív egyszálú RNS vírusok, mint például a Hepatitisz C) virális poliprotein kódoló régiójának transzlációja CAP-független, mivel a picornavírus genom 5’ végéhez nem kapcsolódik CAP, csak egy VPg peptid, amelynek funkciója az RNS védelmét kivéve teljesen eltér a CAP-tól. Továbbá egyes picornavírusok, mint például az enterovírusok a gazdasejtbe bejutva éppen a CAP-függő transzlációhoz szükséges iniciációs faktorokat (pl. eIF-4G) bontanak el, leállítva ezáltal a gazdasejt saját CAP-függő fehérje-gyártó apparátusát. Bővebben lásd a 2014.03.28.-ai bejegyzésemet. A picornavírus 280 és 450 nukleotid hosszúságú IRES régiója komplex másodlagos RNS-struktúrával rendelkezik és számos a gazdasejt által termelt iniciációs faktor (IF) és riboszómális komplex kötőhelyéül szolgál. Az IRES-IF-riboszóma komplex működése, ezáltal a virális poliprotein képződése teljesen CAP-független. Az IRES régió másodlagos RNS struktúrája alapján összesen 5 különböző picornavírus IRES típust (IRES I-V) azonosítottak: Az enterovírus-típusú IRES-I; a parecho-, cardio-, erbo- és aphthovírus típusú IRES-II; a hepatitis A-típusú IRES-III; a Hepacivírus/Pestivírus-típusú IRES-IV valamint a 2012-ben leírt kobu- és salivírus-típusú IRES-V. A felsorolásban szereplő víruscsoportokról, a picornavírusok taxonómiájáról bővebben olvashattok egy későbbi bejegyzésemben. A több doménes (összetett, kompakt struktúra), számos hajtű- és hurok elemből felépülő IRES típusok első ránézésre is eltérő másodlagos RNS-struktúrával rendelkeznek, lásd az alábbi ábrán. A doméneket általában római számokkal, az azokat alkotó hajtű-struktúrákat pedig betűkkel jelölik. A több doménes IRES legfontosabb funkciójú, és struktúrálisan legkonzervatívabb úgynevezett mag-doménjei (a 40S riboszómális alegység kötőhelye) közvetlenül a start kodon (AUG) előtt találhatóak.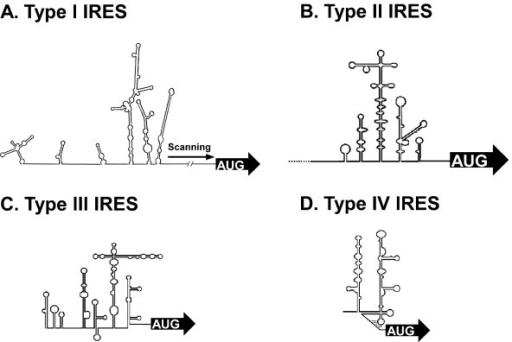
1. ábra: A különböző IRES típusok (I-IV) strukturális jellemzői. AUG = Transzláció iniciációs kodon.
Az eddig ismert emlős picornavírus fajok között a leggyakoribb az ún. IV-es típusú IRES. A 2. ábrán is jól látható, hogy a IV-es IRES mag-doménjeinek mérete, az azokat alkotó bázisok száma között lehetnek kisebb-nagyobb eltérések, de a domének fő szerkezeti vonásai konzervatívak.
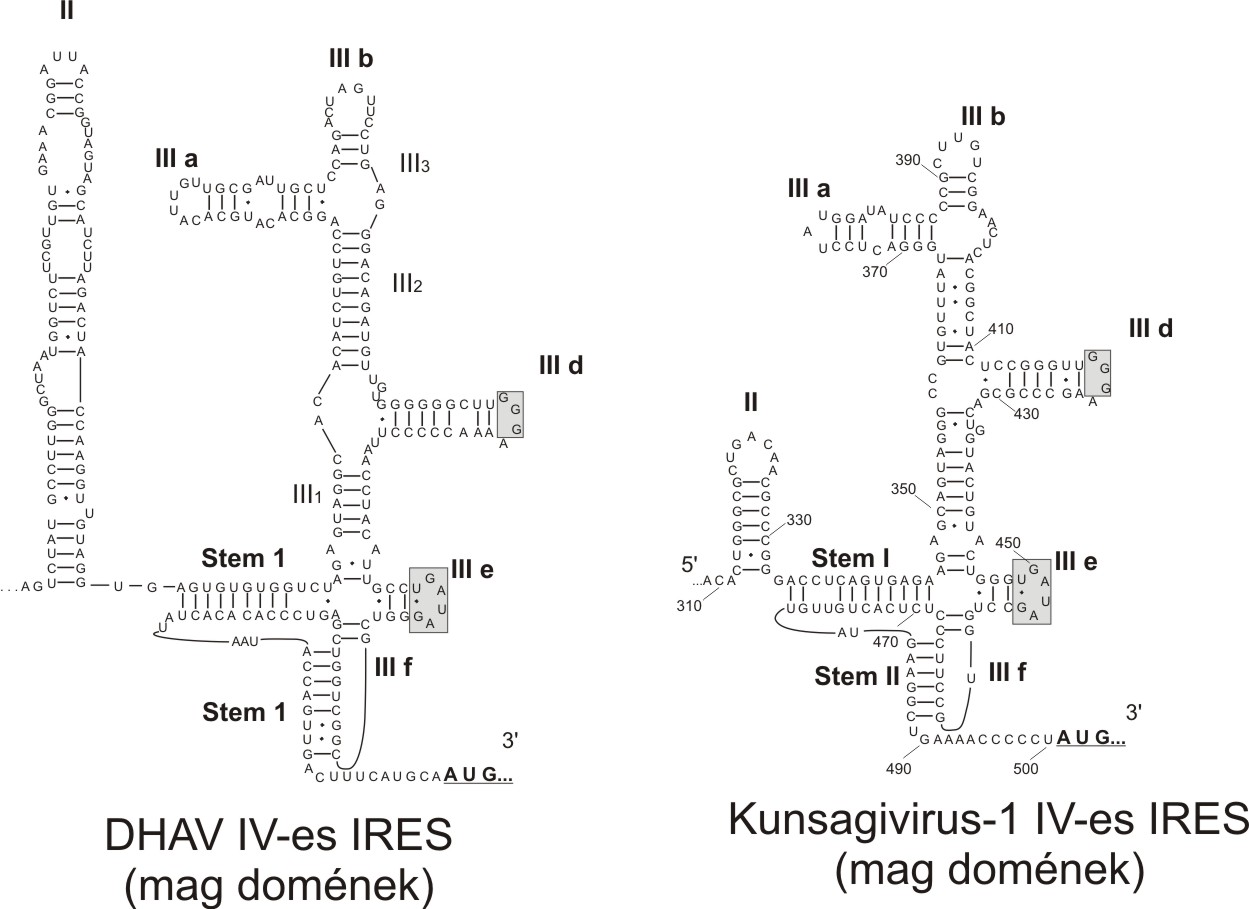
2. ábra: A IV-es típusú IRES mag doménjeinek másodlagos RNS-szerkezete. Az RNS bázisok közötti egyszerű vonalak a kanonikus bázispárokat, a fekete pontok az úgynevezett lötyögős (”wobble”) bázispárokat jelölik. Az egyes domének különböző méretei (pl. a domén II) ellenére a másodlagos szerkezet alapjában véve ugyanolyan a két IRES esetében.
Az emlős picornavírusokhoz hasonlóan az eddig azonosított madár picornavírusok között is a IV-es típusú IRES előfordulása a leggyakoribb. Azonban az emlős IV-es IRES-el ellentétben a legtöbb madár picornavírus IV-es IRES tartalmaz egy kb. 100 nukleotidból álló hajtű-struktúrát, amelynek a csúcsi (apikális) részén egy 20 nukleotidból álló, szekvencia és másodlagos struktúra szintjén is konzervatív, 8-as számot formázó struktúra található (3. ábra). Ez a kb. 100 nukleotidos hajtű általában IV-es IRES III-as doménjének csúcsi részén található, azonban a kacsa hepatitisz A vírusoknál (duck hepatitis A virus = DHAV; Avihepatovirus nemzetség) ez a hosszú hajtű nem a III-as doménen, hanem külön doménként a II-es domén előtt (Ie) található (3. ábra). Eddigi ismereteink szerint a 100 nukleotidos hajtű a 8-as szerű csúcsi résszel a fókákat fertőzi képes Aquamavírus A1 (Aquamavirus nemzetség) kivételével csak a madár picornavírusok jellemzője, és csak a IV-es IRES részeként fordul elő. Az Aquamavírus A1 esetében a DHAV-hoz hasonlóan a 100 nukleotidos hajtű a II. domén előtt található. További érdekesség, hogy nem minden madár picornavírus IV-es típusú IRES-ének képezi részét a fent említett 100 nukleotidos hajtű. Például hiányzik az madár enkefalomielitisz vírus (avian encephalomyelitis virus = AEV; Tremovirus nemzetség); a madár sapelovírus (ASV-1; Sapelovirus nemzetség) és a nemrég leírt Kunsagivírus 1 (”Kunsagivirus” nemzetség) IV-es IRES-eiből. A pontos funkciója a 100 nukleotidos hajtűnek jelenleg ismeretlen, habár a DHAV IRES-en végzett deléciós vizsgálatok kimutatták, hogy az Ie domén (ami megfelel a 100 nukleotidos hajtűnek a 8-as szerű csúcsi résszel) struktúrájának megváltoztatása 40%-al csökkentette az IRES aktivitását. További részletekért lásd Pan és mtsai, 2011: J. Virol. 2012, 86(2):1129. DOI: 10.1128/JVI.00306-11. Mindenesetre a 100 nukleotidos hajtű-mentes IV-es IRES-ek jelenléte az emlős és egyes madár picornavírusok között a hosszú hajtű fontos, de nem-esszenciális funkciójára utalhatnak.
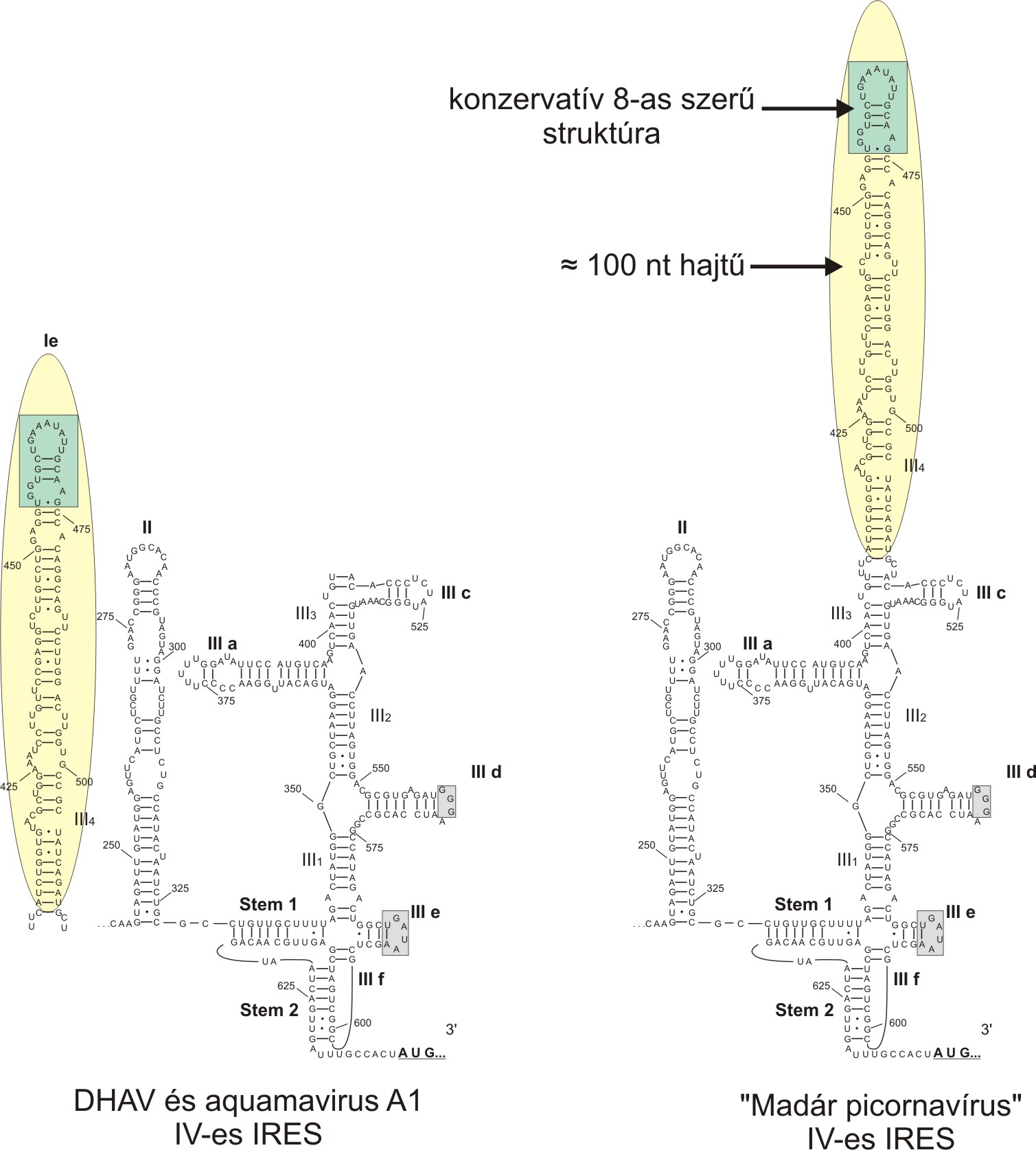
3. ábra: IV-es IRES madár picornavírus változatának másodlagos struktúrája. A 100 nukleotidos (nt) hajtű (sárga háttér) a konzervatív a 8-as szerű csúcsi résszel (zöld háttér) két különböző pozícióban helyezkedhet el.
Referenciák:
Pankovics, P., Boros, A. and Reuter, G., 2012. Novel picornavirus in domesticated common quail (Coturnix coturnix) in Hungary. Arch. Virol. 157, 525-530.
Boros, Á., Kiss, T., Kiss, O., Pankovics, P., Kapusinszky, B., Delwart, E., Reuter, G., 2013. Genetic characterization of a novel picornavirus distantly related to the marine mammal-infecting aquamaviruses in a long-distance migrant bird species, European Roller (Coracias garrulus). J. Gen. Virol. 94, 2029-2035.
Boros, Á., Pankovics, P., Knowles, N.J., Nemes, C., Delwart, E., Reuter, G., 2014. Comparative complete genome analysis of chicken and turkey megriviruses (family Picornaviridae): long 3’ untranslated regions with a potential second open reading frame and evidence for possible recombination. J. Virol. 88(11):6434.DOI: 10.1128/JVI.03807-13.
Ajánlott bejegyzések:
-
 Az első madár kobuvírus leírása - szalakóta kobuvírus 1
Az első madár kobuvírus leírása - szalakóta kobuvírus 1
-
 A súlyzó-szerű struktúra jelenléte két különböző víruscsalád tagjaiban (Picornaviridae és Caliciviridae) - új mozgó genetikai elem jelenléte RNS vírusokban?
A súlyzó-szerű struktúra jelenléte két különböző víruscsalád tagjaiban (Picornaviridae és Caliciviridae) - új mozgó genetikai elem jelenléte RNS vírusokban?
-
 Rendhagyó madár picornavírusok bemutatása 2. rész
Rendhagyó madár picornavírusok bemutatása 2. rész
-
 A nem-strukturális fehérjék funkcióinak áttekintése
A nem-strukturális fehérjék funkcióinak áttekintése
-
 Helyzetjelentés
Helyzetjelentés